白垩纪末植物灭绝事件和现代雨林的起源
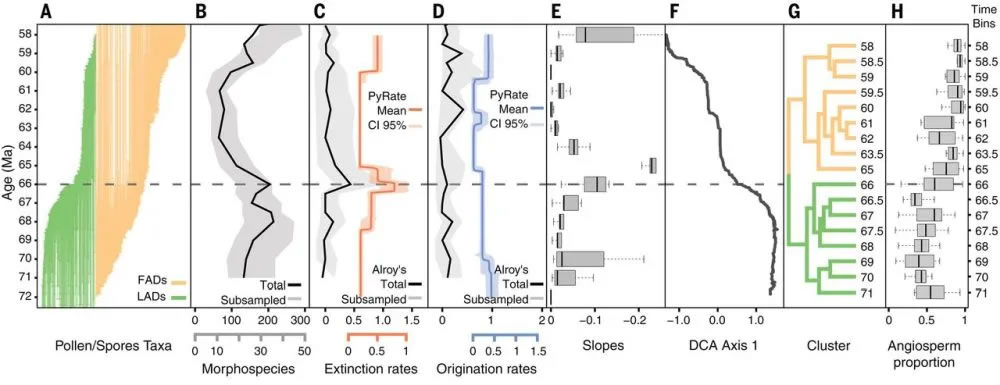
图1南美洲北部马斯特里赫特阶-古新世界限植物多样性和植物组成的变化(Carvalho et al., 2021)。(A)马斯特里赫特阶-古新世界限植物变化(绿色:灭绝植物;桔黄色:新生植物);(B)植物多样性变化;(C)植物灭绝速率;(D)植物产生速率;(E)存活率;(F)植物组成数量;(G)植物组成类型(绿色:马斯特里赫特阶;桔黄色:古新世);(H)被子植物含量

图2 树叶化石形态(Carvalho et al., 2021)
(化石网报道)据中科院地质地球所(撰稿:姜文英/新生代室):发生在白垩纪和古近纪交界(K/Pg界线,~6600万年前)的小行星撞击地球事件,造成了大规模的快速灭绝事件,摧毁了地球上约3/4的生物。南美洲北部距离撞击点Chicxulub大约1500km,坐落在白垩纪-古近纪的古赤道,K/Pg事件对低纬热带雨林的影响仍然不清楚。
巴拿马史密森热带研究所的Mónica Carvalho博士、Carlos Jaramillo教授及其团队最近在Science发表论文(Carvalho et al., 2021)。该团队分析了哥伦比亚K/Pg界限前后5万多粒花粉和6000多块树叶化石,探讨了事件前后低纬热带地区的植物多样性、植物组成和森林结构的变化,以及K/Pg事件在重塑现代热带雨林方面的作用。
一、热带植被的灭绝和反转
南美洲热带的孢粉类型多样性在马斯特里赫特阶(Maastrichtian,~72-66Ma)比古新世中晚期(66-62Ma)高(图1A-图1E),在生物灭绝的高峰期(66Ma),孢粉类型多样性减少了45%,超出了在马斯特里赫特阶或古新世的灭绝量。在K/Pg界限之后,孢粉多样性仍然维持在较低的水平,直到60 Ma之后才基本恢复到灭绝前的水平,之后进一步增加;PETM事件发生时期(~56Ma)甚至超过了灭绝前的数量(Jaramillo et al., 2006, 2010)。
植被组成也在K/Pg界限前后发生了明显的变化(图1F, 图1G),并且这个变化是永久性的。马斯特里赫特阶被子植物和蕨类植物的比例差别不大,分别为47.9%和49.5%,而在古新世被子植物占主导,达84%(图1H)。裸子植物(主要是南洋杉科,Araucariaceae)的含量减少,从马斯特里赫特阶的2.5%,减少为古新世的0.4%。
K/Pg事件影响的范围、生物灭绝的程度以及生态系统恢复的格局,存在明显的地区差异(Jacobs and Currano, 2021)。在巴塔哥尼亚,植物多样性恢复较快,多达90%的植物类型重新出现在丹麦阶(Danian, 66-61.6Ma)(Barreda et al., 2012),而在北美北部大平原,大约30%的植物遭灭绝,植物的恢复较慢,直到古新世末期或始新世早期(~56Ma),植物多样性才恢复到灭绝前的水平(Peppe, 1998;Wing et al., 1995)。
二、叶面形态和森林类型
叶面形态分析的样品来自马斯特里赫特阶Guaduas化石群(2053个化石)、中晚古新世Bogotá化石群和Cerrejón化石群(4898个化石)。
作者在马斯特里赫特阶Guaduas植物化石群识别出41种被子植物和4种蕨类植物,古新世Bogotá植物化石群发现46种被子植物和2种蕨类植物,Cerroejón植物化石群包含58种被子植物、5种蕨类植物和1种针叶树。3个古植物群中被子植物的叶面形态均与现代热带雨林相似,叶子较大,边缘没有锯齿,叶尖细长(图2)。根据叶面积推测,维持Guaduas、Bogotá和Cerroejón古植物群生长需要的年平均降水量分别为234-293cm、182-184cm和240-308cm。
作者比较了Guaduas、Bogotá和Cerroejón等3个古植物群与现代热带雨林树叶的亲缘关系,发现与Guaduas古植物群具有亲缘关系的科一级的被子植物,广泛分布在热带低地,如Lauraceae、Araceae、Theaceae、Arecaceae、Rhamnaceae、Piperaceae、Salicaceae、Canellaceae、Dilleniaceae、Urticaceae和Monimiaceae等。古新世Bogotá古植物群和Cerroejón古植物群相似,则包含了现代热带雨林的主要树种,包括Fabaceae、Euphorbiaceae、Lauraceae、Salicaceae、Violaceae、Malvaceae、Melastomataceae、Rhamnaceae、Arecaceae、Eleaocarpaceae和Araceae等。因此在科一级的植物组成上,古新世古植物群比马斯特里赫特阶古植物群更接近现代雨林。
单位面积叶脉长度的分布(leaf vein length per area: VLA)和叶表皮(leaf cuticles)稳定碳同位素(δ13C)可以反映森林的树冠结构(Canopy structure)。Guaduas古植物群VLA值相对较高,呈单峰分布,树叶δ13C值较低。相反,Cerroejón古植物群VLA值呈双峰分布,树叶δ13C值的范围较大,与郁闭度高、多层结构的现代热带雨林相似。因此,作者推测马斯特里赫特阶的热带雨林,很有可能是较为开放的环境,有利于呼吸作用产生的CO2和大气CO2的混合,林下层和树冠层之间光照强度的垂直差异较小,与多层结构的古新世古植物群相比,这些开阔的森林通过蒸腾作用产生的降雨可能更少。
三、现代雨林的起源
白垩纪以前,热带雨林相对开阔,植被由被子植物,蕨类植物和针叶树(主要是南洋杉科)组成。而古新世森林,则与现代植被非常相似,郁闭度高,多层结构,植被以被子植物为主,其组成在科一级的水平上相似。
马斯特里赫特阶和古新世森林虽然在植物组成和树冠结构方面存在差异,但叶面形态的相似性表明这两个明显不同的生态系统,其气候状况却是相似的,均为湿润的热带气候。不同之处在于马斯特里赫特阶雨林相对开阔,被子植物种类不丰富,针叶树种类尽管很少,但数量相对稳定,另外,碳的固定速率、蒸腾蒸发作用和营养物质的循环可能也比古新世雨林小。而古新世雨林郁闭度的增加,导致光照和水的利用产生明显的垂直差异,为植物新种的出现和生长方式的改变创造了条件,形成了类似现代雨林那样复杂的垂直结构。
这些显著的差异引发2个问题:(1)在白垩纪末,被子植物在种类和生态方面都呈现出多样性,从水生植被到高大树木(Wing and Boucher, 1998; Jud et al., 2018),生长环境多种多样,完全具备形成郁闭树冠的有利条件,可是为何马斯特里赫特阶雨林的郁闭度较低? (2)古新世和马斯特里赫特阶的气候条件十分相似,可是为何古新世雨林没有重新回到与马斯特里赫特阶相似的雨林,反而演变成另外一种不同的植物群落和结构?
由此,作者提出了以下三种现代雨林起源的假设:
(1)大型食草动物的干扰。在撞击事件前,大型食草动物,主要是恐龙,通过不断践踏和大量进食,连续地干扰生境,产生林隙,减少了相邻植物对光照的竞争,从而保持了雨林的开放性。大型食草动物在白垩纪末灭绝,减少了林隙的形成,触发热带植物之间对光的竞争,产生了更多荫蔽的栖息地,导致对光和水分等生长需求不同的新种的产生;
(2)与土壤营养物质有关。马斯特里赫特阶南美洲北部为广袤稳定的低地,经历了持续几百万年的湿润气候,风化作用强烈,导致土壤贫瘠,营养物质较低,只能维持开阔的森林。而撞击事件产生的灰烬丰沃了热带土壤,有利于生长速度比针叶树和蕨类植物快的被子植物的繁殖;
(3)与选择性的灭绝有关。尽管南洋杉科的种类不多,他们是形成晚白垩纪的树冠结构的重要因素,由于南洋杉科的生态范围较窄,以及较少的树木生长方式,极易遭受大规模的灭绝事件。相比之下,马斯特里赫特阶被子植物较高的生态多样性,以及更高的全基因组复制能力,使他们更能抵抗灭绝。因此在白垩纪末,针叶树几乎从热带雨林中消失,为被子植物在古新世期间形成多样化的树冠创造了更多的生存空间。
主要参考文献
Barreda V D, Cúneo N R, Wilf P, et al. Cretaceous/Paleogene floral turnover in Patagonia: drop in diversity, low extinction, and a Classopollis spike[J]. PLoS One, 2012, 7(12): e52455.
Carvalho M R, Jaramillo C, de la Parra F, et al. Extinction at the end-Cretaceous and the origin of modern Neotropical rainforests[J]. Science, 2021, 372(6537): 63-68.
Jacobs B F, Currano E D. The impactful origin of neotropical rainforests[J]. Science, 2021, 372(6537): 28-29.
Jaramillo C, Ochoa D, Contreras L, et al. Effects of rapid global warming at the Paleocene-Eocene boundary on neotropical vegetation[J]. Science, 2010, 330(6006): 957-961.
Jaramillo C, Rueda M J, Mora G. Cenozoic plant diversity in the Neotropics[J]. Science, 2006, 311(5769): 1893-1896.
Jud N A, Michael D D, Williams S A, et al. A new fossil assemblage shows that large angiosperm trees grew in North America by the Turonian (Late Cretaceous)[J]. Science Advances, 2018, 4(9): eaar8568.
Peppe D J. Megafloral change in the early and middle Paleocene in the Williston Basin, North Dakota, USA[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2010, 298(3-4): 224-234.
Wing S L, Alroy J, Hickey L J. Plant and mammal diversity in the Paleocene to early Eocene of the Bighorn Basin[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 1995, 115(1-4): 117-155.
Wing S L, Boucher L D. Ecological aspects of the Cretaceous flowering plant radiation[J]. Annual Review of Earth and Planetary Sciences, 1998, 26(1): 379-421.